TGF-βによって誘導されるマスト細胞のFcεRIとc-Kitの発現低下及び細胞活性化の抑制に転写因子Ehfが関与している
Tweet執筆者情報
執筆者:中野信浩
執筆者所属:順天堂大学大学院 医学研究科 アトピー疾患研究センター
原著論文:The transcription factor Ehf is involved in TGF-β-induced suppression of FcεRI and c-Kit expression and FcεRI-mediated activation in mast cells. (The Journal of Immunology 195:3427-3435, 2015)
更新日:2015年10月14日

概要
免疫抑制及び抗炎症性のサイトカインTGF-β1が抗原刺激によって引き起こされるマスト細胞の活性化を抑制することは以前から知られていたが、その分子的なメカニズムは不明であった。我々はマウスのマスト細胞を用いて、TGF-β1により発現が誘導される転写因子ets homologous factor (Ehf) が、マスト細胞の活性化に関与するいくつかの遺伝子の転写を抑制することでマスト細胞の活性化抑制に寄与していることを明らかにした。
はじめに
マスト細胞は細胞表面に高親和性IgE受容体FcεRIと増殖因子stem cell factor (SCF) の受容体c-Kitを発現している造血幹細胞由来の細胞で、即時型アレルギー反応において中心的な役割を果たしている。FcεRIにはIgEが結合しており、そのIgEに抗原が結合して複数のFcεRIが架橋されると細胞内に活性化シグナルが伝達される。マスト細胞は細胞内にヒスタミン等のケミカルメディエーターを含む顆粒を持っており、活性化に伴い顆粒内の物質を細胞外に放出して、アレルギー疾患の諸症状を引き起こす。
以前から、TGF-β1がマスト細胞の活性化に重要なFcεRIとc-Kitの発現を低下させ、抗原刺激に伴う細胞活性化を抑制することが知られていたが(1-3)、その分子的なメカニズムは不明であった。本研究において、Etsファミリーに属する転写因子Ehfが、TGF-β1によって引き起こされるFcεRIとc-Kitの発現低下及びマスト細胞活性化の抑制に関わっていることを明らかにした。
マスト細胞をTGF-β1で刺激するとEhfの発現が誘導される
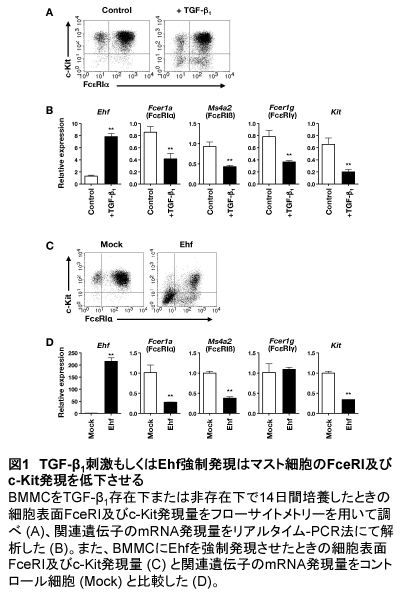
マウス骨髄細胞をIL-3とSCF存在下で分化誘導して得た骨髄由来マスト細胞(BMMC)をTGF-β1存在下で14日間培養すると、細胞表面のFcεRI及びc-Kit発現量が低下した(図1A)。FcεRIは3種類のサブユニットα鎖、β鎖、2つのγ鎖から成る複合体である。TGF-β1存在下で培養したBMMCではFcεRIα、β、γ及びc-Kitをコードしている遺伝子の発現量がコントロール細胞に比べ低下していたことから、FcεRI及びc-Kit発現の低下は転写レベルで制御されていることが示唆された(図1B)。
これらに対し、TGF-β1はEhfの転写レベルを顕著に増加させた(図1B)。Ehfは上皮系細胞で発現していることが知られており、上皮細胞の癌化に関係することが報告されている(4)。また、造血系細胞では樹状細胞の分化に関係することが示唆されている(5)。BMMCにおいて、TGF-βシグナル伝達分子の一つであるSmad4をsiRNAによりノックダウンすると、Ehf遺伝子の発現量増加が見られなくなったことから、Ehfの発現誘導はTGF-β/Smadシグナルに依存していることが示唆された。しかしながら、Smadが直接Ehfのプロモーター領域に結合して転写活性に関与しているかどうかについてはまだ不明である。
マスト細胞にEhfを強制発現させるとFcεRI及びc-Kit発現量が低下する
転写因子Ehfは特定の遺伝子に対して転写抑制因子として働くことが示唆されているが(6)、マスト細胞における役割はまったく不明であったため、BMMCにEhfを強制発現させてその機能を解析することを試みた。レトロウイルスベクターを用いてEhf遺伝子を導入し、これを強制発現させたBMMCでは、TGF-β1非存在下でもコントロール細胞に比べ細胞表面のFcεRIとc-Kit発現量が低下した(図1C)。また、Ehfを強制発現させたBMMCではFcεRIα、FcεRβ、c-Kit遺伝子の転写が抑制されていたが、FcεRIγの転写レベル低下は認められなかった(図1D)。
次に、EhfがFcεRIα、FcεRβ、c-Kit遺伝子の発現を抑制するメカニズムを明らかにするため、ルシフェラーゼ遺伝子の上流に各遺伝子のプロモーター領域の配列を挿入したプラスミドをBMMCに導入し、ルシフェラーゼレポーターアッセイを行って各プロモーターの転写活性を解析した。BMMCにEhfを強制発現させるとFcεRIα、FcεRβ、c-Kit遺伝子のプロモーター活性は顕著に抑制されたが、FcεRIγ遺伝子のプロモーター活性はほとんど影響を受けなかった。この結果は、図1Dの結果を裏付けるものであり、EhfがFcεRIα、FcεRβ、c-Kit遺伝子の転写を直接抑制していることを示唆している。さらに、クロマチン免疫沈降法によりEhfが各プロモーターに結合していることが確認できたので、Ehfはこれらの遺伝子のプロモーター領域に直接結合し、転写抑制因子として働いていると考えられた。
さらに、TGF-β1存在下で培養したBMMCでは、FcεRIα、FcεRβ、c-Kit遺伝子の転写活性を増加させる転写因子をコードする、Spi1(PU.1)、Gata1(GATA-1)、Gata2(GATA-2)遺伝子の転写レベルが低下していた。Ehfを強制発現させたBMMCでは、これらのうちGata1とGata2の転写レベル低下が見られた。EhfがGata1とGata2の転写レベルを低下させる詳細なメカニズムはまだ明らかでないが、FcεRIとc-Kitの発現低下にはこれらの転写活性化因子の転写レベル減少も寄与していると推測される。
EhfはFcεRIの架橋刺激によるマスト細胞の活性化を抑制する
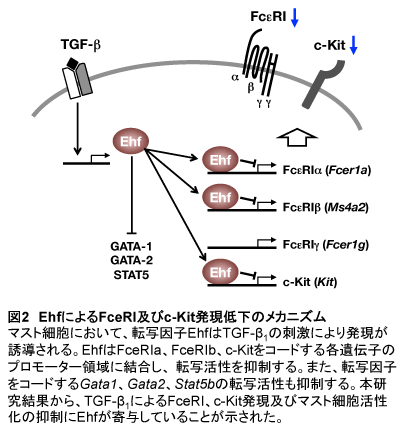
TGF-β1存在下で培養したBMMCとEhfを強制発現させたBMMCは、FcεRIの架橋刺激によって引き起こされる脱顆粒とサイトカイン産生がコントロール細胞に比べ抑制された。脱顆粒の抑制はFcεRIの発現量低下に起因していると考えられるが、サイトカイン産生量の低下はFcεRIの発現量低下だけでは説明できないほど顕著であった。そこで、FcεRIを介してサイトカイン産生を引き起こすシグナル伝達分子について、それらをコードする幾つかの遺伝子の転写レベルを比較した。TGF-β1存在下で培養したBMMC、Ehfを強制発現させたBMMCともにStat5b(STAT5)の転写レベルがコントロール細胞に比べ低下していることを見出した。これらの結果から、EhfはFcεRIα、FcεRβ、c-Kit、STAT5の発現を低下させることでマスト細胞の活性化を抑制している可能性が考えられた(図2)。
おわりに
マウスマスト細胞において、TGF-β1刺激によって発現が誘導される転写因子Ehfを強制的に発現させると、TGF-β1非存在下であってもTGF-β1によって引き起こされるマスト細胞の反応の一部を模倣することができた。これらの結果から、TGF-β1によるマスト細胞のFcεRIとc-Kit発現の低下、及び細胞活性化抑制の一部はEhfによって仲介されていることが示された(図2)。本研究ではin vitroの実験系を用いて細胞内の分子メカニズムの解明を試みたが、生体内でEhfがどの程度重要な働きを担っているのか、またその機能異常が疾患の発症に関わっているかどうか等はまだ明らかになっていない。Ehf欠損マウスはまだ得られていないので、まずはEhf欠損マウスを作製し、本研究で得られた知見を足がかりにその表現型を解析することで、より多くの新規の知見が得られると期待している。
参考文献
1. Gomez G, et al. (2005) J Immunol. 174: 5987-5993.
2. Kashyap M, et al. (2005) Exp Hematol. 33: 1281-1291.
3. Fernando J, et al. (2013) J Immunol. 191: 4505-4513.
4. Stephens DN, et al. (2013) J Biol Chem. 288: 34304-34324.
5. Appel S, et al. (2006) Blood. 107: 3265-3270.
6. Tugores A, et al. (2001) J Biol Chem. 276: 20397-20406.