クロオオアリ触角における化学感覚タンパク質の発現様式と進化
Tweet執筆者情報
執筆者:北條賢1、石井健一2
執筆者所属:1神戸大学 大学院 理学研究科 生物学専攻 2The Salk Institute for Biological Studies
原著論文:Antennal RNA-sequencing analysis reveals evolutionary aspects of chemosensory proteins in the carpenter ant Camponotus japonicus. (Scientific Reports 5:13541, 2015)
更新日:2015年11月24日

概要
社会性昆虫であるアリの、化学物質(匂い)に依存したコミュニケーションでは、触角の化学感覚タンパク質がセンサとして働く。アリは社会の複雑化に伴い、多様なセンサを進化させたと考えられるが、そのレパートリーの全貌は明らかでない。我々はクロオオアリの触角を用いた遺伝子の網羅的解析から、新たに12種の化学感覚タンパク質を発見した。これらセンサにみられた特徴的な発現様式から、アリの社会性進化の謎に迫る。
はじめに
社会性昆虫であるアリは、化学物質を介し綿密な個体間コミュニケーションを行うことが知られています。アリはこの能力を利用し、厳格な分業システムを伴う「コロニー」と呼ばれる組織的な社会を形成します(1)。多くのアリ種において、コロニーは互いに交わらない閉鎖的な性質を示し、働きアリは他コロニーの個体を攻撃的に排除する優れた巣仲間識別能力を持ちます。またアリは巣内でも絶えず個体間コミュニケーションを図り、臨機応変に役割分担をしています。1匹の働きアリの役割(育児、採餌、巣の防衛など)は生まれつき定まっているのではなく、ある限られた巣仲間とだけ交わされる局所的な個体間相互作用を介して、相手が従事する役割を認識し、それに応じて自身の労働を特殊化させることで、巣内の労働分業が成立します。ある個体の労働の特殊化は他個体の行動変化を連鎖的に誘発し、その影響がいずれ自身へもフィードバックされることで、社会として適切な分業体制が自己組織的に形成されていきます。
私たちヒトが皆異なる顔つきをしているのと同様、アリでは体表面に分泌される匂い成分(体表炭化水素)の構成比が個体ごとに少しずつ異なります。アリは触角(昆虫の主要な化学感覚器)を用いて、この「匂いバーコード」の微妙な違いを識別し、様々な個体情報(出身コロニー・役割・繁殖状態・日齢など)を瞬時に読み取った上でコミュニケーションを行うことがわかっています(2)。しかしながら、アリが複雑な匂い成分の混合物からどのように様々な情報を抽出しているのか、その神経分子基盤は未だ明らかにされていません。近年解読が進められた複数種のアリのゲノム情報からは、匂い識別に関わる遺伝子(化学感覚関連遺伝子)のレパートリーが、他の昆虫と比べてアリで特に多様化していることがわかってきました(3)。一方で、それらの遺伝子がアリの化学感覚器においてどのようなパターンで発現し、複雑な感覚受容を担うセンサとしての機能を発揮しているかは、ほとんど明らかになっていませんでした。今回私たちは、クロオオアリ Camponotus japonicus (図1A)の触角からRNAを抽出し、次世代DNAシーケンサおよび大型計算機を用いて発現遺伝子の網羅的解析を行うとともに、触角における化学感覚関連遺伝子の発現部位を詳細に調べました。
化学感覚タンパク質
触角に発現する遺伝子の中で、私たちは化学感覚関連因子の一つである化学感覚タンパク質(Chemosensory protein: CSP)に着目しました。昆虫の触角表面には、触角よりさらに微細な毛状の構造体(化学感覚子)が多数見られ、その内部には、脳へとつながる受容神経細胞が血リンパ液に取り囲まれる形で存在します。「匂いバーコード」を構成する体表炭化水素は水に溶けにくいため、本来であればリンパ液にはじかれてしまい、触角表面から内部の神経細胞まで到達できません。一方、血リンパ液中に存在するCSPは、脂溶性と水溶性の両方を兼ね備えた特殊な構造をもっています。この性質は、CSPが体表の匂いと神経細胞・脳との間の橋渡し役として働くことを可能にします。すなわちCSPは、脂溶性の体表炭化水素と結合し、親水環境であるリンパ液から神経細胞膜上の受容体まで運搬するのです(4)。このお陰で、アリは自身の触角を他のアリ個体の表面に接触させることにより、相手の匂い成分を触角内部に染み込ませ、脳に情報を伝えてその先の行動を決めることが出来るようになります。
先行研究により、CSPはアリ触角に発現するタンパク質の大部分を占める主要因子であることが明らかとなっていました(5,6)。また他の昆虫におけるCSPの解析例からは、CSPに構造の異なる複数のタイプが存在することが知られ、化学物質への結合特性に差があると予想されていました。
アリは社会生活を営まない他の昆虫と比較して、より複雑なコミュニケーションを要するため、進化の過程で匂いセンサ(CSP)の豊富なレパートリーを獲得している可能性が考えられます。また、アリ触角表面に存在する微細な化学感覚子にも、形状やサイズ、数のバリエーションがみられます。
これら複数種のCSPと化学感覚子の存在からは、両者をうまく組み合わせる(各CSPが特定の化学感覚子に発現する)ことにより、多彩な化学コミュニケーションが可能になると予想できます。しかしながら、アリのCSPのバリエーションがどのくらいあるのか、また触角においてどのような発現パターンや機能を示すのか、全体像の理解には至っていませんでした。私たちは、アリがなぜこのような高度な社会性を成立させることができたか、その進化の謎を解く鍵がCSPにあると考え、研究に着手しました。
私たちの研究グループではこれまでに、クロオオアリの主要なCSPの一つをCjapCSP1と名付け、その機能を解析してきました。その結果、i) CjapCSP1がアリの体表炭化水素と結合すること、ii) その結合が神経細胞の活性化を導くこと、iii) 体表炭化水素の由来(同じ巣のアリか、異なる巣のアリか)により神経興奮の度合いに差が生じることを、世界に先駆けて報告してきました (6)。一方で、CjapCSP1に結合しない体表炭化水素も多数存在すると予想されたことから、この因子だけではアリの個体間コミュニケーションの仕組みを説明しきれないとも考えました。そこで今回、クロオオアリがもつCSPのレパートリーを網羅的に調べることを目標に、採取したアリの触角からRNAを抽出し、次世代シーケンサによる遺伝子解析を行いました。クロオオアリのゲノム情報は未だ公表されていませんが、他のアリや昆虫のゲノム情報も参考に、得られた配列情報からさらなるCSPの探索を行い、12個のクロオオアリCSP遺伝子(CjapCSP1〜10, 12, 13)を見出すことに成功しました。

CSP遺伝子の進化
次に私たちは、アリのCSPの進化的背景を探ることを目的に、昆虫種間での比較を試みました。CjapCSPのアミノ酸配列情報と様々な昆虫の化学感覚タンパク質の配列情報を用いて、最尤法による分子系統解析を行いました。その結果、前述のCjapCSP1から進化的に派生したと考えられる3つのCSP (CjapCSP10, 12, 13)が、アリ類で特別に多様化したCSPであることがわかりました(図1B)。
では、配列が少しずつ異なるこれらのCSPは、進化の過程でどのような選択を受けてきたのでしょうか。CSP遺伝子間で塩基配列を比較し、同義置換(アミノ酸配列の変化を導かない差異)より非同義置換(アミノ酸配列の変化を導く差異)が多くみられれば、非同義置換によるタンパク質の性質変化が進化上有利に働いて残された(正の自然選択を受けた)と考えられます。
各CSPの塩基配列から非同義置換と同義置換との比率(dN/dS比)を算出したところ、アリ特異的に多様化したCSPのうち、CjapCSP12, 13では正の自然選択(dN/dS>1)が検出されました。この結果は、CSPが、アリの生存に役立つ性質をもって適応的な進化を遂げたことを示唆するものです。
働きアリにおけるCSPの発現部位と触角でのカースト特異性
各化学感覚タンパク質の働きを推測する上では「身体のどの部位で、どれくらいの量が発現しているか」という情報が重要な手掛かりとなります。私たちは働きアリの身体の各部位からRNAを抽出し、定量PCRにより発現量を算出して比較しました。その結果、化学感覚器に集中して発現しているのは、12個のCjapCSPのうち4個だけであると判明しました。前述のCjapCSP12, 13は、炭化水素を運搬するCjapCSP1とともにこの中に含まれており、触角を利用したコミュニケーションに関わる化学感覚タンパク質だと推測されます。この「全てのCjapCSPが化学感覚器に発現しているわけではない」という結果を別の見方で解釈すると、これまで知られていた「触角での匂い分子の橋渡し」という役割以外にも、CSPが身体の各所で未知の機能を発揮している可能性が考えられます。
アリの社会では、カースト(メスの働きアリ・メスの次世代女王・オス)間でコミュニケーション様式に差がみられます(例えば、オスは巣仲間認識が全くできない)。興味深いことに、触角で検出された先ほどのCjapCSP 1, 12, 13は、カースト間で発現量に違いがみられました。このことから、これら3つの触角CSPが、アリのカースト特異的な機能を担っている可能性が考えられます。
触角におけるCSPの発現様式
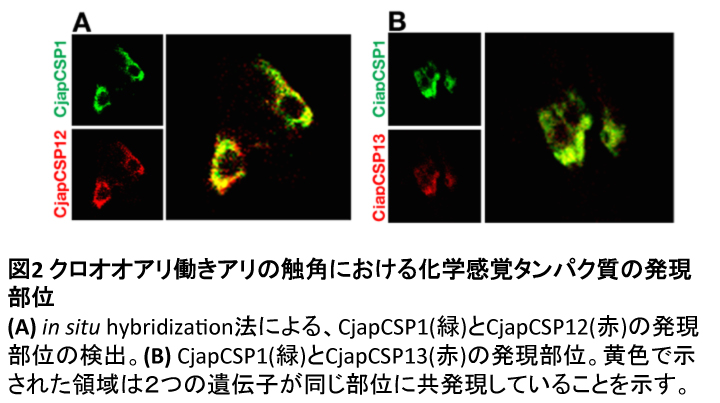
アリの体表炭化水素は、触角表面の微細な毛状器官の一つである錐状感覚子で受容され、CjapCSP1もこの感覚子で主に発現しています(6)。また上記の発現量解析から、CjapCSP12, 13が1と同様に、触角で多く発現していることがわかりました。そこで、今回新たに見出したCjapCSP12, 13がこの感覚子で発現しているかを、in situ hybridization法を用いて組織学的に解析しました。すると、両者が触角内でCjapCSP1と同じ部位に検出され、これらが体表炭化水素を受容する感覚子で共発現していることが示唆されました (図2)。アリ類で特異的に進化した化学感覚タンパク質どうしが、触角上の同一の感覚子で協力して働くことにより、複雑な個体間コミュニケーションが可能になっていると考えられます。
おわりに
今回私たちは、アリの主要な化学感覚タンパク質であるCSPのレパートリーを解明し、それらの進化と機能について考察を深めてきました。アリで特別に多様化したCSPは、匂い成分の混合物が織りなす複雑なバーコードから、様々な個体情報を読み取り高度な社会を築く上で、重要な役割を担うことが示唆されました。本研究は、アリ社会でみられる洗練されたコミュニケーションの分子基盤やその進化の理解に貢献すると期待されます。各CSPのさらなる機能解析により今後、アリがどのように複雑な社会組織を成立させたか、という謎に挑戦していきたいと、私たちは考えています。
著者ウェブサイト
■ 北條賢
個人ページ:https://sites.google.com/site/hojokmasaru/
■ 石井健一
researchmap:http://researchmap.jp/ken1ishi2/
ResearchGate:https://www.researchgate.net/profile/Kenichi_Ishii2/publications
所属研究室:http://asahina.salk.edu/personnel.php
参考文献
1. Holldobler B & Wilson EO (1990) The Ants, Belknap Press.
2. VanderMeer RK et al. (1998) Pheromone communication in social insects, Westview Press.
3. LeBoeuf AC et al. (2013) Curr Opin Neurobiol 23:3-10.
4. Pelosi P et al. (2006) Cell Mol Life Sci 63:1658-1676.
5. Ishida Y et al (2002) Naturwissenshaften 89:505-507.
6. Ozaki M et al (2005) Science 309:311-314.