晝惈堚揱偡傞怘帠惈旍枮
Tweet幏昅幰忣曬
幏昅幰丗怷揷弮戙丄杧嫃戱榊丄栘懞旤崄丄峳堜桬擇丄婽堜峃晉丄彫愳壚岹丄敤揷弌曚
楢棈愭丗敤揷弌曚丄仹371-8512孮攏導慜嫶巗徍榓挰3-39-15丂孮攏戝妛 惗懱挷愡尋媶強僎僲儉壢妛儕僜乕僗暘栰丂嫵庼丂hatada@gunma-u.ac.jp
尨挊榑暥丗Paternal Allele Influences High Fat Diet-Induced Obesity. PLoS One 9: e85477, 2014.
峏怴擔丗2014擭1寧24擔

奣梫
C57BL/6J (B6)儅僂僗偼崅帀朾怘傪怘傋偝偣傞偙偲偵傛傝旍枮偵側傝傗偡偄偑丄偦偺堚揱揑攚宨偵偮偄偰偼傢偐偭偰偄側偄丅崱夞変乆偼偙偺怘帠桿摫惈偺旍枮偑晝恊偐傜偺傒堚揱偡傞偙偲偲丄偙偺尰徾偑晝惈僀儞僾儕儞僩堚揱巕偺Igf2, Peg3偺敪尰偲娭學偟偰偄傞偙偲傪曬崘偡傞丅
偼偠傔偵
歁擕椶偵偍偄偰晝偲曣桼棃偺僎僲儉偼摍壙偱側偄丅偙傟偼僎僲儉僀儞僾儕儞僥傿儞僌乮僎僲儉嶞傝崬傒乯偑懚嵼偟丄僀儞僾儕儞僩堚揱巕偲偄偆晝恊桼棃傾儗儖摿堎揑偁傞偄偼曣恊桼棃傾儗儖摿堎揑側敪尰傪帵偡堚揱巕偑偁傞偙偲偵傛傞丅僀儞僾儕儞僩堚揱巕偺拞偵偼旍枮偵娭學偡傞傕偺偑懡偔抦傜傟偰偍傝丄椺偊偽Prader-Willi徢岓孮偲偄偆旍枮傪敽偆幘姵偺愑擟堚揱巕偼僀儞僾儕儞僩堚揱巕偱偁傞(Butle, 2011)丅傑偨Peg1丄Dlk1丄Peg3偲偄偭偨堚揱巕傕旍枮偲娭學偟偰偄傞丅偦偺懠丄帀朾偺暘壔偲娭學偟偰偄傞傕偺傕抦傜傟偰偄傞丅
C57BL/6J (B6)儅僂僗偼崅帀朾怘偵傛傝旍枮傗俀宆偺摐擜昦偵側傝傗偡偔(Surwit et al., 1988) 丄挿擭戙幱尋媶偵梡偄傜傟偰偒偨丅AKR/J傗DBA/2J儅僂僗傕摨條偵怘帠桿摫惈旍枮偵側傝傗偡偄偑丄偦傟偵懳偟偰A/J 丄KsJ丄 SWR/J偼怘帠桿摫惈旍枮偵側傝偵偔偄丅崱夞偺尋媶偱偼丄PWK儅僂僗偑怘帠桿摫惈旍枮偵側傝偵偔偄偙偲傪尒偄弌偟偨丅偦偟偰怘帠桿摫惈旍枮偵側傝傗偡偄B6偲PWK傪岎屳偵岎攝偟偰(PWK亊B6) F1偲(B6亊PWK) F1偺俀庬椶偺F1傪偮偔傝旍枮偲僀儞僾儕儞僥傿儞僌偺娭學偵偮偄偰挷傋偨丅
怘帠桿摫惈旍枮偼晝惈堚揱偡傞
C57BL/6J (B6)儅僂僗偼怘帠惈桿摫旍枮偵側傝傗偡偔丄PWK儅僂僗偼怘帠桿摫摫惈偺旍枮偵側傝偵偔偄丅偙傟傜偺儅僂僗傪偐偗崌傢偣偰F1傪偮偔偭偨偲偙傠B6偑晝恊偺偲偒丄偡側傢偪(PWK亊B6) F1偼怘帠惈桿摫旍枮偵側傝傗偡偄偑丄PWK偑晝恊偺偲偒偼丄偡側傢偪(B6亊PWK) F1偼怘帠惈桿摫旍枮偵側傝偵偔偐偭偨(恾1A)丅偮傑傝怘帠惈桿摫旍枮偼晝惈堚揱偡傞偙偲偑傢偐偭偨丅傑偨B6儅僂僗偼懴摐擻堎忢傪帵偡偑丄摨偠傛偆偵F1挷傋偰傒偨偲偙傠B6偑晝恊偺偲偒偺傒偱懴摐擻堎忢傪帵偟丄PWK偑晝恊偺偲偒偼懴摐擻堎忢偑傒傜傟側偐偭偨丅偡側傢偪懴摐擻堎忢傕晝惈堚揱偡傞偙偲偑傢偐偭偨(恾1B)丅
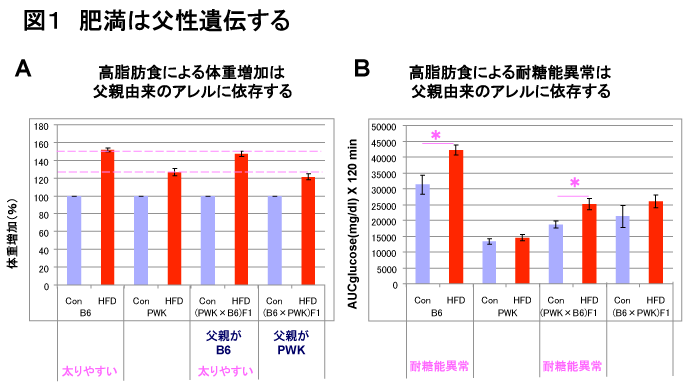
怘帠惈旍枮偵傛傝敀怓帀朾偺僀儞僾儕儞僩堚揱巕偺敪尰偑曄壔偡傞
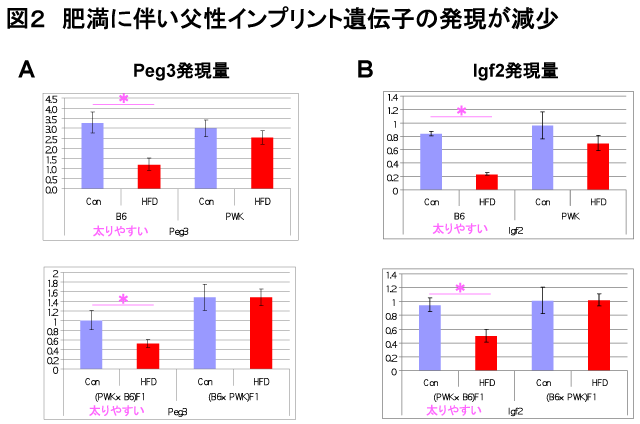
僀儞僾儕儞僥傿儞僌偲偼堚揱巕敪尰偑桼棃偡傞恊偑晝偱偁傞偐曣偱偁傞偐偵傛傝敪尰偑堎側傞尰徾傪偄偆丅晝惈僀儞僾儕儞僩堚揱巕偼晝恊桼棃偺傾儗儖偺傒敪尰偟丄曣惈僀儞僾儕儞僩堚揱巕偼曣恊桼棃偺傾儗儖偺傒敪尰偡傞丅偙傟傜偺堚揱巕偺拞偵偼旍枮傗帀朾偺暘壔偵娭學偟偰偄傞傕偺傕懡偔抦傜傟偰偄傞丅偦偙偱晝惈堚揱偡傞怘帠惈桿摫旍枮偑僀儞僾儕儞僩堚揱巕偺敪尰偲娭學偟偰偄傞偲峫偊丄B6偲PWK偺敀怓帀朾偵偍偗傞僀儞僾儕儞僩堚揱巕偺敪尰傪挷傋偨丅偦偺寢壥丄晝惈敪尰堚揱巕偺Peg3偲 Igf2偺敪尰偑崅帀朾怘傪怘傋偝偣傞偙偲偵傛傝B6偱偺傒敪尰偑尭彮偟偰偄偨(恾2)丅傑偨曣惈敪尰堚揱巕偺Igf2r偺敪尰偑B6偱偺傒崅帀朾怘傪怘傋偝偣傞偙偲偵傛傝敪尰偑忋徃偟偰偄偨丅偝傜偵嫽枴怺偄偙偲偵F1偺敀怓帀朾偱敪尰傪挷傋偨偲偙傠B6偑晝恊偺偲偒丄偡側傢偪(PWK亊B6) F1偼Peg3偲Igf2偺敪尰偑崅帀朾怘傪怘傋偝偣傞偙偲偵傛傝尭彮偡傞偑丄PWK偑晝恊偺偲偒丄偡側傢偪(B6亊PWK) F1偼Peg3偲Igf2偺敪尰偑尭彮偟側偐偭偨(恾2)丅偡側傢偪怘帠惈旍枮偺晝惈堚揱偵敽偭偰僀儞僾儕儞僩堚揱巕偺敪尰偑曄壔偡傞偙偲偑傢偐偭偨丅曣惈敪尰堚揱巕偺Igf2r偼F1偱敪尰偺曄壔偼側偐偭偨丅
Peg3偲Igf2偺DNA儊僠儖壔偼曄壔偑側偄
僀儞僾儕儞僩堚揱巕偱偼晝曣偺傾儗儖偱DNA儊僠儖壔偑堎側傞椞堟偑偁傝Differentially Methylated Region (DMR)偲屇傇偑丄偙傟偼傾儗儖摿堎揑側敪尰偲娭楢偟偰偄傞丅偦偙偱崅帀朾怘傪怘傋偝偣傞偙偲偵傛傝偍偙偭偨敪尰偺曄壔偵偼丄DNA儊僠儖壔偺曄壔偑娭梌偟偰偄傞偺偱偼側偄偐偲峫偊敀怓帀朾偱偺DNA儊僠儖壔忬懺傪挷傋偨丅偟偐偟側偑傜DNA儊僠儖壔偵偼曄壔偑側偔丄敪尰偺曄壔偼懠偺婡峔偵傛傞傕偺偲峫偊傜傟傞丅
Igf2偼TNF-alpha偵傛傞墛徢嶌梡偐傜敀怓帀朾傪庣偭偰偄傞
嬤擭丄旍枮偼敀怓帀朾偺枬惈墛徢偲枾愙偵娭楢偟偰偄傞偙偲偑傢偐偭偰偒偨(Cinti et al., 2005)丅帀朾慻怐傊偺儅僋儘僼傽乕僕偺廤愊偼懱廳偺憹壛丄帀朾偺墛徢丄僀儞僗儕儞姶庴惈偺掅壓偲娭楢偟偰偄傞丅旍枮偟偨儅僂僗偺敀怓帀朾偱偼墛徢惈働儌僇僀儞偱偁傞MCP-1偺暘斿偑憹偊丄儅僋儘僼傽乕僕偑梀憱偟偰偒偰丄墛徢惈偺僒僀僩僇僀儞偱偁傞TNF-alpha偺暘斿傪懀恑偡傞丅偦傟偵傛傝帀朾嵶朎偺墛徢偼憹埆偟帀朾偺僀儞僗儕儞姶庴惈偺掅壓偑傕偨傜偝傟傞(De Taeye et al., 2007)丅偝傜偵MCP-1偺暘斿偑憹偊傞偲偲傕偵丄僀儞僗儕儞庴梕懱偺姶庴惈傪椙偔偡傞傾僨傿億僱僋僠儞(Ravussin, 2002)偺暘斿偑尭彮偡傞丅
変乆偼Igf2偑帀朾嵶朎偵偳偺傛偆側塭嬁傪梌偊傞偐傪敀怓帀朾嵶朎偺儌僨儖偱偁傞3T3-L1嵶朎偱挷傋偨丅偦偺寢壥Igf2偺揧壛偵傛傝TNF-alpha偵傛傞MCP-1偺敪尰憹壛丄傾僨傿億僱僋僠儞傗僌儖僐乕僗僩儔儞僗億乕僞乕偱偁傞Glut4偺敪尰尭彮傪梷偊傞偙偲偑偱偒偨丅偝傜偵Igf2偼TNF-alpha偵傛傞僀儞僗儕儞埶懚揑側摐庢傝崬傒偺尭彮傕梷偊傞偙偲偑偱偒偨丅埲忋偺偙偲偐傜Igf2偼偦偺峈墛徢嶌梡偵傛傝旍枮傪梷惂偟偰偄傞偙偲偑帵嵈偝傟偨丅偙偺傛偆側岠壥偼Peg3偱偼傒傜傟側偐偭偨丅
Discussion
崱夞偺尋媶偱怘帠桿摫惈旍枮姶庴惈偺B6偲懴惈偺PWK傪偐偗偁傢偣偨偲偙傠F1偱偼B6偑晝恊偺偲偒偺傒怘帠桿摫惈旍枮偵側偭偨丅偙偺偙偲偐傜怘帠桿摫惈旍枮偼晝惈堚揱偟僀儞僾儕儞僩堚揱巕偺娭梌偑悇應偝傟偨丅偦偙偱旍枮偟偨儅僂僗偺敀怓帀朾偵偍偗傞僀儞僾儕儞僩堚揱巕偺敪尰傪挷傋偨偲偙傠晝惈敪尰偺僀儞僾儕儞僩堚揱巕Peg3偲 Igf2偺敪尰偑尭彮偟偰偄偨丅偙偺尰徾偑偍偙傞尨場偲偟偰偼乮侾乯偙傟傜偺堚揱巕偺揮幨挷愡椞堟偺B6偲PWK偺懡宆偵傛傞偺偐丄乮俀乯偙傟傜偺堚揱巕偺敪尰傪惂屼偡傞晝惈敪尰偺僀儞僾儕儞僩堚揱巕偺B6偲PWK偺懡宆偵傛傞側偳偑峫偊傜傟傞丅偝偰丄偙傟傜偺堚揱巕偺敪尰尭彮偑旍枮偵娭學偡傞偐偱偁傞偑丄Peg3偼僲僢僋傾僂僩偡傞偲懱帀朾検偑憹壛偡傞偙偲偑曬崘偝傟偰偄傞(Curley et al., 2005)丅Igf2偼IGF僔僌僫儖傪惂屼偡傞堚揱巕偱偁傝丄偦偺敪尰偺尭彮偑帀朾検偺憹壛偲娭學偡傞偙偲偑儅僂僗偱曬崘偝傟偰偍傝(Jones et al., 2001)丄僸僩偱寣拞偺IGF2偑彮側偄偲旍枮偺儕僗僋偑憹偡偲偄偆曬崘偑偁傞(Sandhu et al., 2003)丅傑偨僸僩偺IGF2偺懡宆偲旍枮偲偺娭楢惈偵偮偄偰偺偄偔偮偐偺曬崘偑偁傞(O'Dell et al, 1999; Gaunt et al, 2001)丅
敀怓帀朾偵偍偗傞Igf2偺敪尰掅壓偼枬惈墛徢傪偲偍偟偰旍枮傪傂偒偍偙偟偰偄傞偐傕偟傟側偄丅崅帀朾怘傪怘傋偝偣偨B6偺敀怓帀朾偱偼MCP-1偺敪尰偑忋徃偟丄Glut4傗傾僨傿億僱僋僠儞偺敪尰偼尭彮偟偰偄傞丅MCP-1偼墛徢惈偺僒僀僩僇僀儞偱偁傞TNF-兛側偳偺暘斿傪懀偟丄敀怓帀朾偺婡擻晄慡傪傕偨傜偡(De Taeye et al. 2007)丅傑偨傾僨傿億僱僋僠儞偼峈墛徢惈偺僒僀僩僇僀儞偱偁傝僀儞僗儕儞姶庴惈傪槾恑偟(Ravussin, 2002)丄寣拞擹搙偺掅壓偼俀宆摐擜昦偲枾愙偵娭楢偟偰偄傞(Matsuzawa, 2010)丅変乆偼Igf2偑TNF-alpha偵傛傞MCP-1偺忋徃偲Glut4偲傾僨傿億僱僋僠儞尭彮傪梷偊傞偙偲傪尒偄弌偟偨丅偙偺偙偲偼Igf2偑峈墛徢嶌梡偵傛傝摐戙幱傪夵慞偟偰偄傞偙偲傪帵嵈偡傞丅
寢榑偲偟偰B6儅僂僗偺崅帀朾怘偵傛傞怘帠桿摫惈偺旍枮偲懴摐擻堎忢偼晝惈堚揱偟丄偦傟偼僀儞僾儕儞僩堚揱巕偺Peg3偲Igf2偺敪尰偑敀怓帀朾偱尭彮偡傞偙偲偲娭學偟偰偄偨丅偙偺偙偲偼Peg3偲Igf2偑旍枮偲摐戙幱傪夵慞偡傞偙偲傪帵嵈偡傞丅傑偨幚嵺偵3T3-L1帀朾嵶朎傪巊偭偨幚尡偱Igf2偼峈墛徢嶌梡傪帩偪丄摐戙幱傪夵慞偟偰偄偨丅
嶲峫暥專
- Butler MG (2011) Prader-Willi Syndrome: Obesity due to Genomic Imprinting. Curr Genomics. 12:204-215.
- Cinti S, Mitchell G, Barbatelli G, Murano I, Ceresi E, Faloia E, Wang S, Fortier M, Greenberg AS, Obin MS (2005) Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J Lipid Res 46:2347-2355.
- Curley JP, Pinnock SB, Dickson SL, Thresher R, Miyoshi N, Surani MA, Keverne EB. FASEB J (2005) Increased body fat in mice with a targeted mutation of the paternally expressed imprinted gene Peg3. FASEB J 19:1302-1304.
- Gaunt TR, Cooper JA, Miller GJ, Day IN, O'Dell SD (2001) Positive associations between single nucleotide polymorphisms in the IGF2 gene region and body mass index in adult males. Hum Mol Genet 10:1491-1501.
- Jones BK, Levorse J, Tilghman SM (2001) Deletion of a nuclease-sensitive region between the Igf2 and H19 genes leads to Igf2 misregulation and increased adiposity. Hum Mol Genet 10:807-814.
- Matsuzawa Y (2010) Adiponectin: a key player in obesity related disorders. Curr Pharm Des 16:1896-1901.
- O'Dell SD, Bujac SR, Miller GJ, Day IN (1999) Associations of IGF2 ApaI RFLP and INS VNTR class I allele size with obesity. Eur J Hum Genet 7:821-827.
- Ravussin E (2002) Adiponectin enhances insulin action by decreasing ectopic fat deposition. Pharmacogenomics J 2:4-7.
- Sandhu MS, Gibson JM, Heald AH, Dunger DB, Wareham NJ (2003) Low circulating IGF-II concentrations predict weight gain and obesity in humans. Diabetes 52:1403-1408.
- Surwit RS, Kuhn CM, Cochrane C, McCubbin JA, Feinglos MN (1988) Diet-induced type II diabetes in C57BL/6J mice. Diabetes 37:1163-1167.