脳特異的DISC1結合タンパク質DBZはオリゴデンドロサイトの分化を促進する
Tweet執筆者情報
執筆者:清水尚子、小山佳久、伊藤彰、遠山正彌
執筆者所属:近畿大学 東洋医学研究所 分子脳科学研究部門 〒589-8511 大阪府大阪狭山市大野東377-2
原著論文:DBZ, a CNS-specific DISC1 binding protein, positively regulates oligodendrocyte differentiation. Glia. in press, 2014. (PMID: 24481677)
更新日:2014年3月7日

概要
統合失調症発症の分子機序は未だ明らかにされていない。我々は、精神疾患多発家系から発見された脆弱性因子DISC1の結合タンパク質であるDBZ (DISC1-binding Zinc finger protein)に着目し、その機能を解析した。DBZはオリゴデンドロサイトに発現しており、ミエリン形成時期である生後14日をピークに発現上昇した。またDBZノックアウトマウスを用いた解析から、DBZはオリゴデンドロサイトの分化を促進することを明らかにした。
諸言
統合失調症は人口の1%に見られ、思春期から青年期にかけて発症する慢性化しやすい精神疾患である。妄想、幻覚や統制を欠いた行動、興奮などの陽性症状と、会話・思考内容の貧困化や社会的ひきこもり等の陰性症状に大別される重大な精神症状が見られる。DISC1遺伝子はスコットランドの精神疾患多発家系(統合失調症、躁鬱病など)を対象とした遺伝学的研究により発見された染色体転座の部位にコードされる遺伝子である1,2。この遺伝子が転座により分断されることがDISC1タンパク質の異常を引き起こし、精神疾患発症のリスクを高めると考えられている。そこで我々はDISC1の転座により分断される部位に結合する分子の結合が不可能となり、その正常な機能を失うことで脳の発達障害が生じるのではないかと想定し、転座部位に結合する分子の探索を開始した。その結果、DISC1の結合タンパク質としてDBZ (DISC1-binding Zinc finger protein)と名付けた新規因子を同定した。
近年、DBZがニューロンの軸索の周りに存在する髄鞘(ミエリン)を形成するオリゴデンドロサイトで機能している可能性を示唆する論文が報告された3。オリゴデンドロサイトの分化異常を示すOlig1ノックアウトマウスのマイクロアレイ解析で、オリゴデンドロサイトの成熟化に関わる数多くの遺伝子とともにDBZの発現が低下していることが報告されている。
統合失調症患者死後脳の研究でオリゴデンドロサイトに発現する因子の発現低下がみられることや、オリゴデンドロサイトの細胞数の減少、配置異常、ミエリンの形成異常がみられることから精神疾患発症に及ぼすオリゴデンドロサイトの役割が重要であると考えられている4,5。これらのことから精神疾患発症に関わる分子メカニズムの解明するために、オリゴデンドロサイトにおけるDBZの機能解析を行った。
DBZの発現検討
in situ ハイブリダイゼーションの結果、マウス脳梁におけるDBZ mRNAの発現は、生後ミエリン形成が盛んな時期(生後0日〜生後3週間)に一過的に発現増加する(図1 A,B)。脳梁におけるDBZ mRNAの発現は生後14日にピークに達し、その後adultにかけて発現減少する(図1 C)。この発現様式はミエリン構成因子であるMBP mRNAの発現と一致した(図1D)。 脳梁RNAによる定量リアルタイムPCRでも生後14日にDBZ、MBP mRNAの発現ピークがあることを確認した(図1E,F)。また、DBZが発現している細胞がオリゴデンドロサイトかどうかを確認するため、in situ ハイブリダイゼーションと免疫染色によって、オリゴデンドロサイト特異的なOlig2陽性細胞にDBZが発現していることを確認した(図1G)。オリゴデンドロサイトの分化のどの時期にDBZが発現しているか検討するため、オリゴデンドロサイト前駆細胞に発現するPDGFRと、それ以降に発現するCC1の抗体を用いてそれぞれDBZ mRNAの発現を検討したところ、DBZ陽性細胞中の約3割がPDGFR陽性、約7割がCC1陽性であった(図1G)。ウエスタンブロット法によりMBPタンパク質は生後14日からadultにかけて発現増加するのに対し、DBZ タンパク質は生後14日に発現が一過的に増加することを見出した(図1H)。
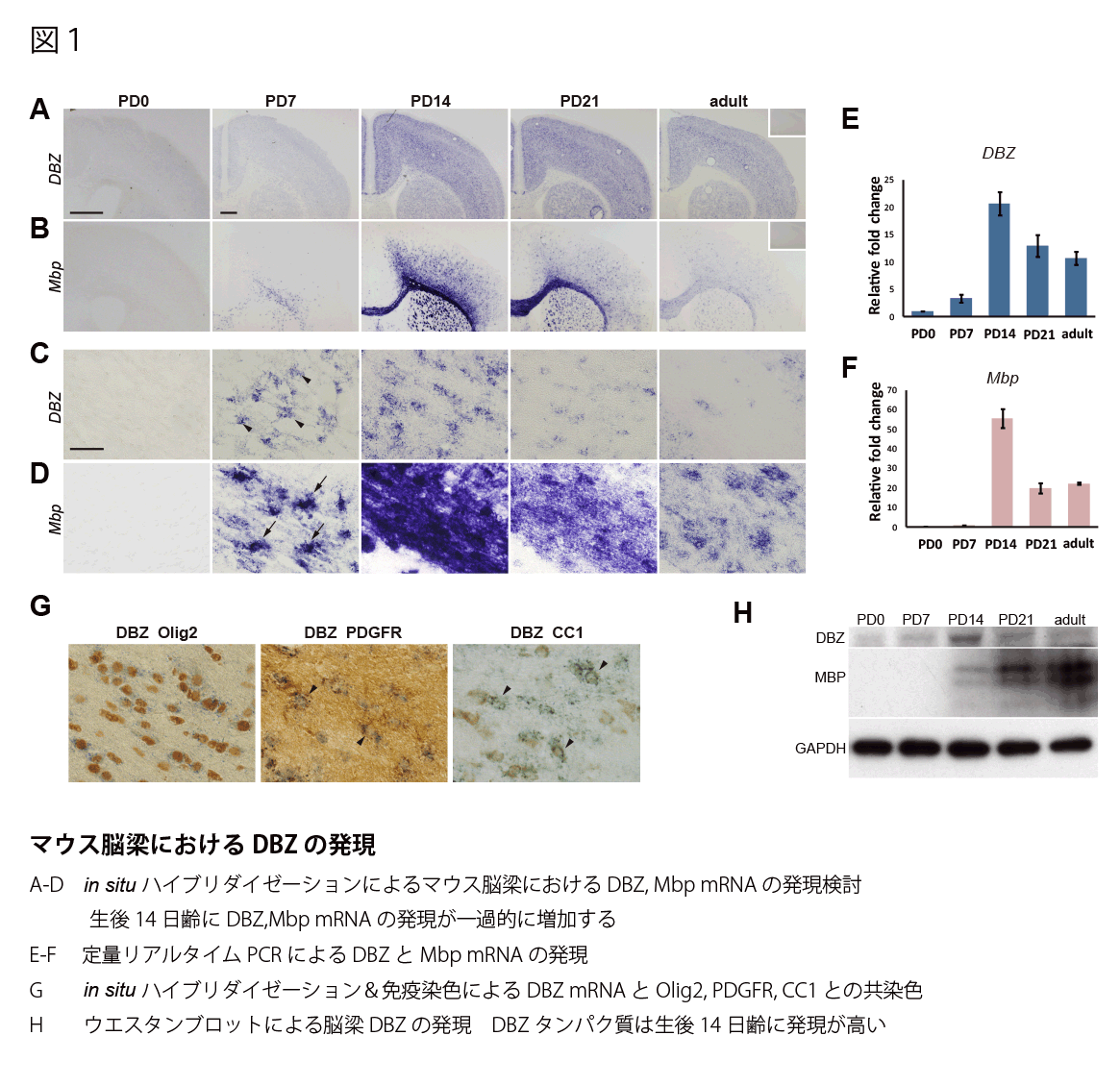
DBZノックアウトマウスの解析
DBZノックアウトマウスがオリゴデンドロサイトの分化異常を示すかどうかを知るために、生後10日齢と成体マウスのミエリン関連因子の発現をin situ ハイブリダイゼーションと免疫染色によって検討した。その結果、生後10日齢のDBZノックアウトマウスでMBP mRNAの発現減少がみられた(図2A,C)。また免疫染色でも明らかにDBZノックアウトマウスのMBPやMAGの発現が低下していた(図2E,G)。一方、成体マウスになるとMAG、MBPのミエリン関連因子の発現の差は、野生型マウスと比較してDBZノックアウトマウスで見られなかった(図2B,D,F,H)。電子顕微鏡でのミエリンの形態観察を行ったところ、生後10日齢ではDBZノックアウトマウスではミエリンの形成が著しく減少していたが、成体マウスでは変化がみられなかった(図2I,J)。脳梁RNAによる定量リアルタイムPCRでも生後10日齢のDBZノックアウトではMAG、MBPの発現減少がみられ、成体では差がなかった(図2K,L)。脳梁のウエスタンブロットによる検討でも生後10日齢のDBZノックアウトマウスではMBPの発現減少がみられ、成体では差がなかった(図2M)
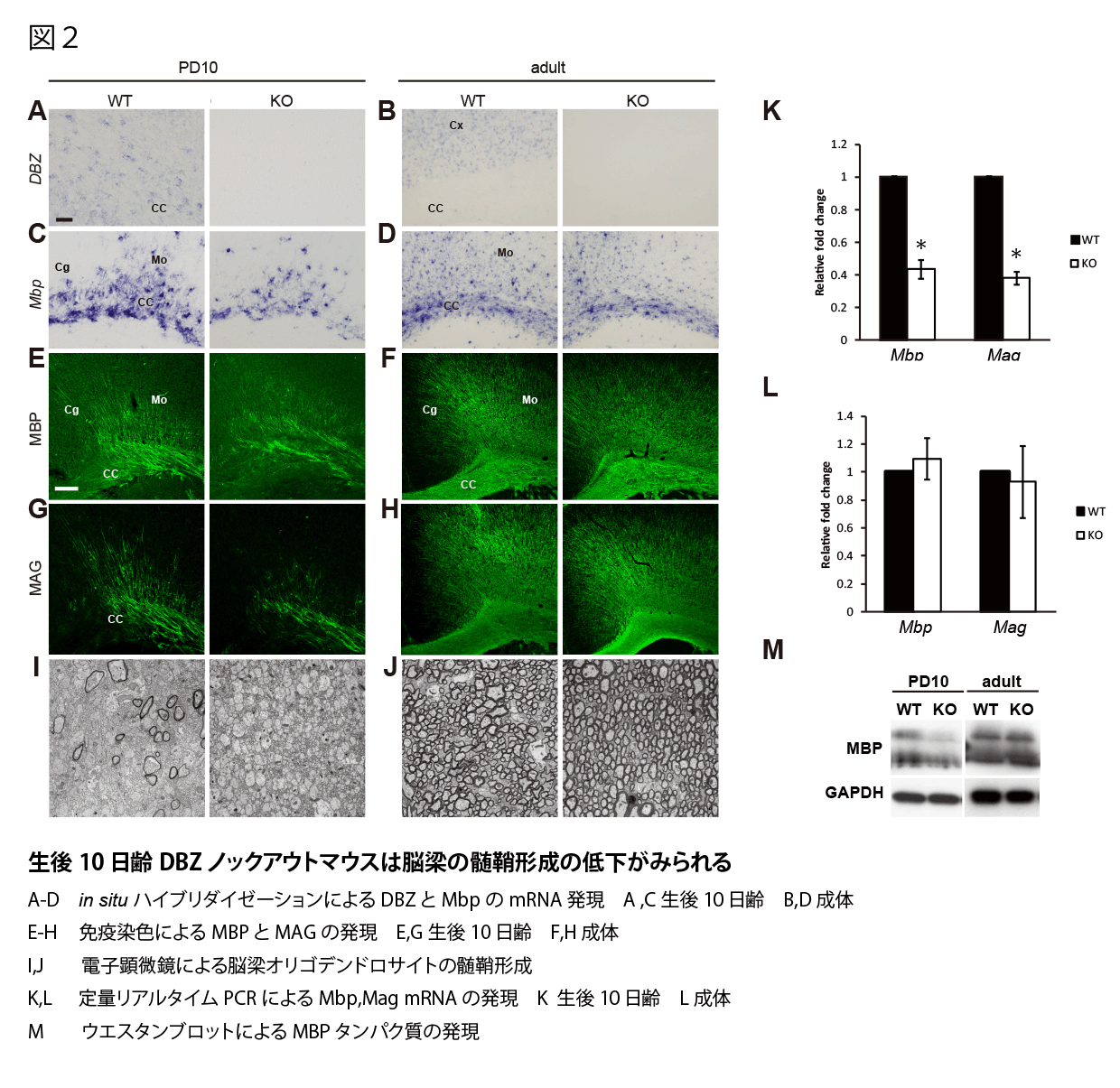
オリゴデンドロサイト初代培養の解析
ラットオリゴデンドロサイト初代培養にPDFGとbFGFを添加すると、オリゴデンドロサイトの増殖が起こる。この条件下で、DBZの発現をsiRNAにより抑制するとオリゴデンドロサイトの増殖はコントロールと比較して差がみられないことから、DBZはオリゴデンドロサイトの増殖には影響を与えないことがわかった。またPDGF(−)、bFGF(−)の条件下ではオリゴデンドロサイトの分化が起こる。分化誘導時にDBZの発現が一過的に増加することが明らかとなった。またこの条件下でDBZの発現を抑制するとMAG、MBP、CNPaseの発現が減少し、オリゴデンドロサイトの分化が抑制された。また、DBZを発現抑制するとオリゴデンドロサイトの分化を負に制御する転写因子であるHes5, Id2, Id4, Tcf4の発現増加がみられ、また正に制御する転写因子Nkx2.2の発現が減少した。以上の結果から、DBZはオリゴデンドロサイトの分化を促進する因子であることを見出した。
考察
DBZはオリゴデンドロサイトに発現しており、生後ミエリン形成のピーク時に発現が一過的に増加する。生後の髄鞘形成が盛んな時期である生後10日齢におけるDBZノックアウトマウスの検討では、ミエリン関連因子の発現が有意に減少していた。また、電子顕微鏡による脳梁のニューロンの軸索の周囲のミエリンがDBZノックアウトマウスではほとんど形成されていないことがわかった。しかし、成体になるとミエリン関連因子の発現、形態に差がみられない。このことから幼若期のオリゴデンドロサイトの分化、ミエリン形成にDBZが必要であるが、その機能は成体では相補されてしまい、差がみられなくなったと思われる。最近の論文でも、オリゴデンドロサイト特異的にdominant active なβ-catenin 発現マウスのマイクロアレイ解析で、オリゴデンドロサイトの成熟化に関わる遺伝子の発現減少がみられ、その中にDBZの発現低下も報告された6。これらのことからもDBZがオリゴデンドロサイトの分化に関わることが強く示唆された。
参考文献
1. Millar JK et al. (2000) Hum Mol Genet 9:1415-1423.
2. Blackwood DHR et al. (2001) Am J Hum Genet 69:428-433.
3. Chen Y et al. (2009) NatNeurosci 11:1398-1408.
4. Edgar N and Sibille E. (2012) Trasl Psychatry 2:e109.
5. Fields R. (2008) Trends Neurosci 31:361-370.
6. Fancy SPJ et al. (2011) Nat Neurosci 8:1009-1018.